Удвоение генов, ответственных за формирование головы, предопределило эволюционный успех позвоночных
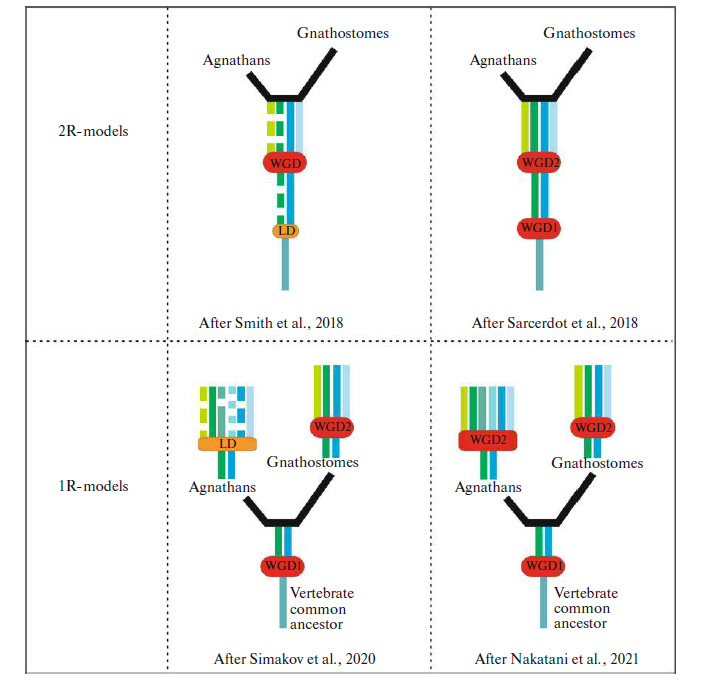

Уже давно известно, что в ходе эволюции позвоночных животных несколько раз происходило удвоение генома. Это установлено, в частности, по присутствию в геноме современных позвоночных многочисленных паралогов — генов-двойников, произошедших от дупликации предкового гена и в дальнейшем эволюционировавших независимо друг от друга. Все позвоночные прошли как минимум два раунда дупликации, а у костных рыб был еще один. Несмотря на то, что мы знаем о количестве удвоений в различных эволюционных группах, хронология этих удвоений и функции образовавшихся паралогов во многом остаются неисследованными, в частности из-за сравнительно небольшого количества данных по геномам некоторых групп позвоночных. Именно этими группами и занялись российские ученые, в своем масштабном исследовании проанализировавшие семейства генов Noggin и Foxg1, ответственных за формирование передних отделов мозга и, видимо, предопределяющих план строения позвоночных в целом.
Полногеномная дупликация — процесс, при котором в клетке удваиваются все хромосомы (и весь содержащийся на них генетический материал). Он неоднократно происходил в самых разнообразных группах организмов — например, у мечехвостов, позвоночных и даже у одноклеточных дрожжей. Наиболее вероятно, что полногеномная дупликация стала стабильным инструментом эволюции из-за ее мгновенной выгодности — возможности создания копий одного и того же гена, способных эволюционировать независимо. Например, у позвоночных наблюдается дупликация генов группы Hox, ответственных за формирование продольной оси тела, и генов, играющих роль в формировании нервного гребня. Клетки-потомки нервного гребня обнаруживаются в самых разных органах: сердце, нервной системе, зубах, костях черепа, а также среди пигментных клеток. Дупликация увеличивает количество эволюционных путей, по которым может пойти организм, что отчетливо видно на примере костистых рыб. У них был «лишний» раунд дупликации генома по сравнению с остальными позвоночными, и они являются самой разнообразной группой позвоночных (из 65 тысяч видов позвоночных около 30 тысяч представлены костистыми рыбами), что, возможно, является результатом увеличения количества «сырого материала» для эволюции.
После полногеномной дупликации далеко не всегда вторая копия генома сохраняется полностью: она может частично утрачиваться (хотя бывают и исключения, например, у шпорцевой лягушки сохраняется тетраплоидный геном). Изначально дупликация возникает в результате нарушения деления клетки. Это может случиться или во время возникновения у родительской особи половой клетки, или на ранних стадиях развития зародыша — на этапе, когда хромосомы должны разойтись в две дочерние клетки, деление останавливается, и в одной клетке остаются четыре копии каждой хромосомы, поэтому все потомки такой клетки имеют в два раза больше хромосом.
Это состояние нестабильно — часто у тетраплоидных клеток нарушается процесс деления, у потомков их носителей теряются или возникают новые участки хромосом, что может повлечь болезни (включая онкологические) и раннюю смерть потомков. С течением времени потомки такой клетки «пытаются» вернуться к исходному диплоидному состоянию. Часть копий хромосом уничтожается, а в части начинают накапливаться изменения, благодаря чему при делении они перестают восприниматься как копии друг друга — именно такие копии и являются следами дупликации, остающимися в эволюционной истории. Гены, прошедшие дупликацию, можно обнаружить не только по их непосредственной схожести, но и по их соседям, которые часто сохраняют схожесть и последовательность на хромосоме, которая наблюдалась до дупликации.
Именно так, вероятно, образовались семейства генов Noggin и Foxg1, ставшие предметом изучения российских ученых, статья которых была недавно опубликована в Paleontological Journal. В данной статье были исследованы «не модельные», но важные для понимания эволюции позвоночных организмы: миноги, хрящевые рыбы и хрящевые ганоиды (то есть осетры и их родственники). Их крайне сложно содержать в лабораторных условиях (в отличие от более привычных модельных организмов), поэтому по ним существует сравнительно мало исследований. Тем не менее, их важно изучать для формирования полноценной картины эволюции позвоночных, так как все они представляют поворотные моменты в эволюции этого подтипа.

Рис. 2. Филогенетическое дерево современных позвоночных. Группы, исследованные в обсуждаемой публикации, отмечены красным. Рисунок из обсуждаемой статьи
Как и у всех позвоночных, у миног (Petromyzontiformes) имеется разделенный на отделы головной мозг, спинной мозг, жаберные дуги. Они обладают высокой способностью к регенерации и адаптивной иммунной системой (возможно, возникшей независимо от челюстноротых позвоночных). Однако от остальных позвоночных их отличает отсутствие челюстей, наличие шишковидной железы (превратившейся в мозге челюстных в шишковидное тело), два полукруглых канала во внутреннем ухе и отсутствие парных конечностей. Одна из особенностей миног на генетическом уровне — так называемый «миножий диалект»: по неизвестной причине часто у неродственных групп миног возникают схожие мутации в одних и тех же генах, из-за чего при анализе отдельных генов возникает иллюзия близких родственных связей между неродственными видами миног и отсутствия родственных связей между генами миног и других позвоночных там, где они должны быть. Из-за этого при изучении геномов миног необходимо корректировать расчеты, чтобы достоверно определить их родство друг с другом и другими позвоночными.
Хрящевые рыбы (Chondrichthyes) — акулы, скаты и химеры. Это самая базальная (самая близкая к эволюционному предку) группа среди челюстных, уникальными чертами которой являются наличие хрящевого скелета и отсутствие плавательного пузыря. Их чешуя гомологична зубам позвоночных: и те, и те состоят из дентина, а внутрь чешуек акул, как и в зубы других позвоночных, заходят кровеносные сосуды. Кроме того, и зубы, и чешуя акул происходят из клеток нервного гребня. Это очень древний признак, утерянный у большинства рыб, но сохранившийся у хрящевых. Общий предок хрящевых и современных костных рыб жил 450 миллионов лет назад. Из-за того, что разделение этих линий произошло так давно, хрящевые рыбы очень важны для понимания эволюции ключевых черт челюстных — парных плавников, мозжечка, жаберного аппарата и, конечно же, челюсти. Однако содержать и исследовать их в лабораторных условиях крайне трудно — им необходим объемный морской аквариум, они откладывают мало икры и икра эта скрыта за непрозрачной оболочкой, что осложняет исследование зародышей и определение стадии развития без вскрытия икринки.
Хрящевые ганоиды (или хрящекостные, Chondrostei) отделились от общего древа развития позвоночных 345 миллионов лет назад и включают в себя веслоносов и осетров — «живых ископаемых», мало изменивших свой внешний облик за миллионы лет. На молекулярном уровне они обладают уникальными свойствами — ядра их клеток содержат огромное число хромосом, от 120 до 380, и среди этих хромосом не выделяются половые, что позволяет геному беспрепятственно удваиваться большое число раз. В отличие от костистых рыб, в хромосомах хрящевых ганоидов часто происходят делеции (мутации, при которых часть генома теряется), из-за которых исчезают единомоментно крупные куски хромосом или даже целые хромосомы. Этот механизм, предположительно, поддерживает стабильность клеток ганоидов.
Ранее упомянутые гены семейств Noggin и Foxg1 присутствуют у всех этих групп (как и у всех хордовых в принципе), однако их количество различается у разных групп. Эти гены ответственны за формирование разнообразных морфологических структур — в частности, уникального строения головы позвоночных. Обнаружение гомологичных генов (или ортологичных, то есть возникших в ходе видообразования у двух разных видов, имеющих общего предка, — в отличие от паралогичных генов, появившихся в геноме одного вида как следствие дупликации) этих семейств у челюстных и бесчелюстных станет подтверждением того, что второй раунд полногеномной дупликации произошел у их общего предка, а не независимо.

Рис. 3. Представители «не модельных» таксонов, ставших предметом изучения в обсуждаемой статье. Речная минога (Lampetra fluviatilis, бесчелюстные ), большая белая акула (Carcharodon carcharias, хрящевые рыбы), белуга (Huso huso, хрящевые ганоиды). Фотографии с сайта ru.wikipedia.org
Семейство генов Noggin (название произошло от английского сленгового слова со значением «голова») в эмбриональном развитии ответственно за формирование у зародыша дорсальной и вентральной (спинной и брюшной) сторон. Если заблокировать у эмбриона развитие спинной стороны и ввести ему белковые продукты Noggin, у него запустится формирование спины. Однако на этом функция этого семейства генов не заканчивается: оно участвует в формировании хрящевой ткани, слухового аппарата, глаз, фолликул волос, также продукты гена Noggin способны подавлять развитие раковых клеток.
Авторы проанализировали геномные базы данных и обнаружили, что у беспозвоночных хордовых (ланцетники и асцидии) в геноме присутствует всего одна копия гена Noggin, в то время как у всех позвоночных имеется несколько паралогов этого гена. Копии этого гена у миног оказались гомологичны таким же копиям у других позвоночных, однако «миножий диалект» затрудняет проведение прямой гомологии между этими копиями кроме одной — NogginD у миног и Noggin4 у других позвоночных. Аналогичная ситуация возникает и при анализе соседних генов: общая картина у челюстных и бесчелюстных совпадает, однако с полной уверенностью установить гомологию нельзя. Кроме того, было обнаружено, что ген Noggin1 отсутствует у всех хрящевых рыб, кроме самых примитивных химер, а у тех акул, у которых его обнаружили, он потерял возможность синтезировать белок. Вероятно, это произошло из-за изменения строения их хрящевого скелета, благодаря чему надобность в этом гене отпала (в опытах над другими позвоночными он подавлял развитие хрящевой ткани).
Таким образом, гены семейства Noggin играют у позвоночных важную роль в эволюции скелета, а также участвуют в формировании мозга, что подтверждается окрашиванием in situ (рис. 4). В эмбрион вводится меченая краской комплементарная (совместимая) копия РНК специфического белка, она соединяется с РНК, ответственной за создание целевого белка в тех клетках, в которых он синтезируется, а из клеток, в которых белок не синтезируется — вымывается. Таким образом, становится видно, в каких участках клетка создает целевой белок, и это позволяет предполагать, какую функцию он несет в организме. Например, у зародышей миног белки-продукты генов семейства Noggin сосредотачиваются в различных районах мозга и хорды, из чего можно сделать вывод, что он играет роль в формировании головного мозга и скелета.

Рис. 4. Экспрессия генов семейства Noggin в зародыше речной миноги, визуализированная окрашиванием продуктов этих генов in situ; стадия роста головы. Рисунок из обсуждаемой статьи
Одной из важнейших особенностей позвоночных является деление их мозга на отделы, среди которых выделяется передний мозг, отсутствующий у всех других групп животных. На поздних стадиях эмбрионального развития передний мозг разделяется на промежуточный мозг и конечный мозг, отвечающий за зрение, движение, формирование рефлексов, а также высшей нервной деятельности. Именно за формирование переднего мозга ответственно второе изученное в этой работе семейство генов — foxg1.
Нарушение работы генов этого семейства приводит к нарушению развития мозга, у человека — к эпилепсии и микроцефалии. Этот ген (или его гомологи) выявлен у нескольких групп беспозвоночных, однако у них он наблюдается всего в одной копии, в то время как авторы выявили три паралога foxg1 у бесчелюстных рыб (для которых до этого была описана всего одна копия гена), также они установили присутствие трех гомологов foxg1 у хрящевых рыб, хрящевых ганоидов и костных рыб, в то время как у млекопитающих и птиц эти гомологи были утеряны, у них осталось всего одна копия foxg1.
Как и в случае с генами Noggin, у бесчелюстных трудно установить однозначное сходство их паралогов foxg1 с генами этого же семейства у челюстных рыб. Поэтому нельзя однозначно утверждать, что их гены гомологичны, несмотря на присутствие схожих соседей этих генов у бесчелюстных и челюстноротых. Окрашивание продуктов foxg1 в зародышах миног и осетров показало, что этот ген экспрессируется в различных областях переднего мозга и верхней губе зародыша.

Рис. 5. Филогенетическое дерево белков гена foxg1 у позвоночных, построенное по методу максимального правдоподобия. Рисунок из обсуждаемой статьи
Таким образом, это исследование показывает, что уникальная особенность позвоночных — «новая голова» — появилась в результате нескольких раундов полногеномной дупликации с дальнейшим сохранением отдельных семейств генов, в том числе ответственных за формирование головы. Эти гены многофункциональны и отвечают не только за строение головы, но и формирование многих других клеток и тканей. В частности, ген noggin1 ответственен за баланс между хрящевой и костной тканью у позвоночных. Изменение интенсивности его работы или даже его утеря приводят к масштабным перестройкам в строении не только головы, но и всего тела, включая скелет, не говоря уже о том, что на ранних зародышевых стадиях это семейство определяет формирование спинной и брюшной стороны.
Авторам, к сожалению, не удалось установить точную эволюционную историю полногеномных дупликаций — несмотря на свидетельства в пользу двух раундов удвоения у общего предка бесчелюстных и челюстных (таких как, например, сходные соседние гены). Из-за особенностей геномов миног (тот самый «диалект») не удается достоверно установить, гомологичны ли удвоенные копии генов Noggin и foxg1 у них генам челюстных позвоночных. Данная работа проливает свет на малоизученные эволюционные линии и показывает, что в их геномах таится еще много вопросов, решение которых поможет полнее понять эволюционную историю формирования позвоночных.
Владимир Вержбицкий
Рис. 1. Предлагаемые сценарии полногеномной дупликации у позвоночных; представлены два основных типа моделей. Согласно первому, две дупликации произошли у общего предка челюстноротых и бесчелюстных. Согласно второму, у него произошел только один раунд дупликации, а второй возник независимо у обеих групп. WGD — полногеномные дупликации, LD — локальные дупликации. Рисунок из обсуждаемой статьи